Multiple meanings: Neural manifolds are inspired by the rich mathematical field of topology, but the meaning of the term in neuroscience has evolved beyond its mathematical origins.
The diversity of behaviors across the animal kingdom is staggering, from feats of agility to nuanced social interactions. Ultimately, these complex behaviors arise from the chatter of neurons, and cracking their “code” has driven systems neuroscience for decades. Indeed, since the 1960s, two observations have captivated neuroscientists. First, the performance of even simple tasks, such as looking at a flash of light, evokes activity in many neighboring neurons. Is this merely built-in redundancy, or does it hint at a more distributed code? Second, individual neurons respond to large numbers of features, such as the color and shape of visual stimuli. Does this “mixed selectivity” reflect ingrained properties of neural function, or is it epiphenomenal?
A body of recent work has suggested a parsimonious explanation for these observations: Perhaps information is not uniquely encoded in the spike trains of individual neurons but rather specified by the coordinated activity of neural populations. By analogy, each of the individual words in this article has meaning, but the information therein becomes clear only when considering all the words together.
Over the past decade, a growing number of my colleagues in the field have begun to formalize a neural manifold framework to elaborate this population-centric view. Indeed, the use of the term “neural manifolds” has become common in neuroscience. These neural manifolds are inspired by the rich mathematical field of topology—in which a manifold is defined as a locally Euclidean topological space.
Despite the rising popularity of neural manifolds, a key question remains: Are they the latest jargony buzzword or a pathway toward wholly new insights into brain function? Polarized debate regularly erupts on social media, and an increasing number of papers try to argue for and against the utility of neural manifolds. I believe many apparent disagreements come from a lack of clarity on what the term really encompasses. Here, I hope to clarify some misconceptions and advocate for neural manifolds as not only a useful analytical framework, but also a conceptually appropriate level at which systems neuroscientists can study the brain.
W
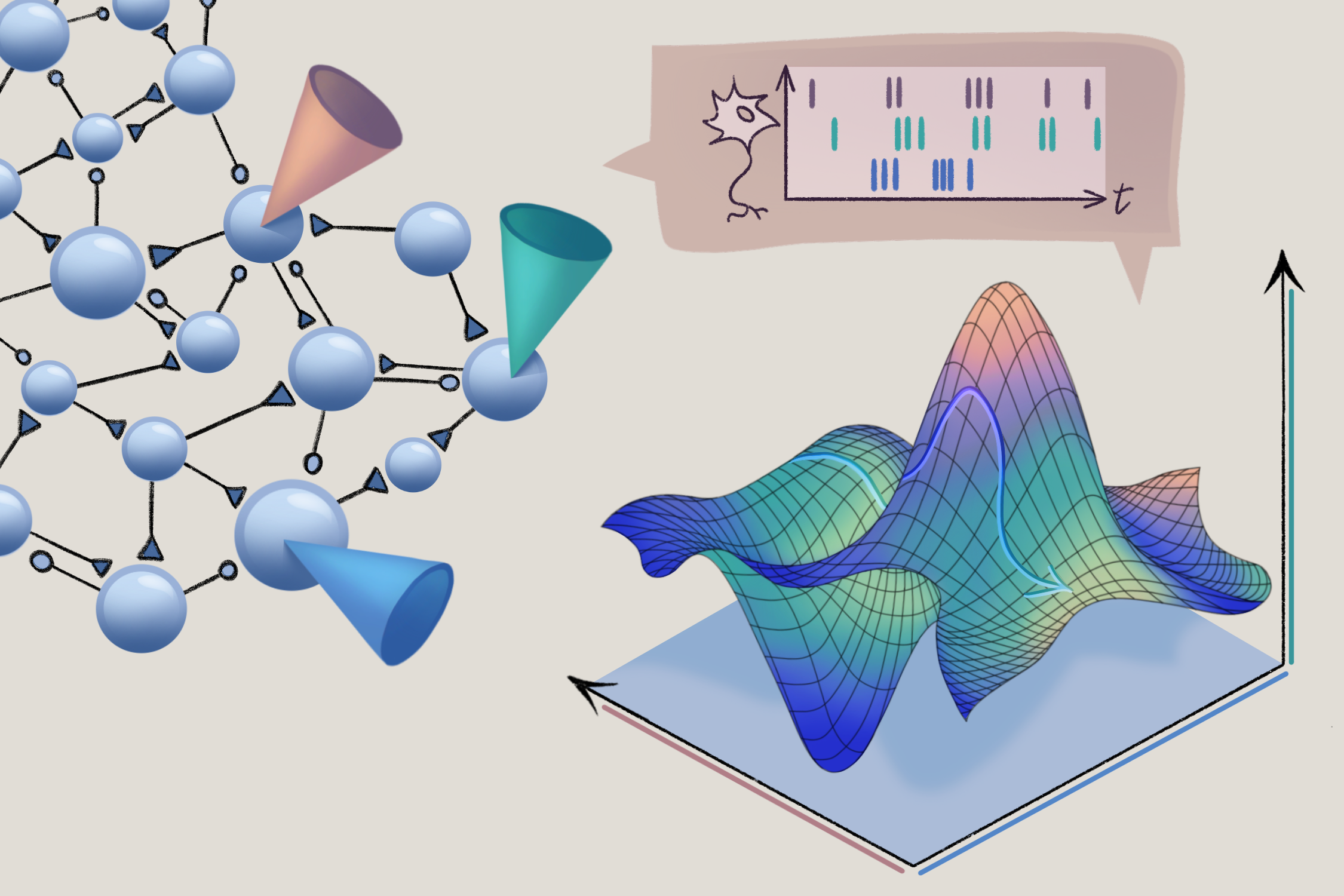
hat is a neural manifold, anyway? To conceptualize the neural manifold, imagine a space where each axis represents the activity of a neuron (the figure below shows a three-neuron example). During some behavior, such as reaching for a coffee cup, each individual neuron’s activity changes, tracing out a trajectory of neural activity in the state space. In many species and brain regions, neural trajectories inhabit considerably fewer dimensions (or degrees of freedom) than the number of neurons. An early demonstration of putative neural manifolds dates to 2003, when Mark Stopfer and his colleagues characterized olfactory encoding in the locust, and the approach has since become ubiquitous throughout systems neuroscience in multiple brain regions and species.
Peaks and valleys: Neural manifolds capture constraints on the activity of groups of neurons. To visualize manifolds, researchers plot firing rates estimated from neural recordings in a space in which each axis represents an individual cell. A 3D representation of three neurons is shown here.
These observations underpinned the neural manifold framework, yet the question remained: Are manifolds convenient visualizations or clues to deeper phenomena? A breakthrough came when Sadtler et al. used a brain-computer interface learning paradigm—in which experimenters can precisely control the mapping from neural activity to behavior—to causally demonstrate that neural manifolds constrain how monkeys can modulate neural population activity. Since then, many studies have gone deeper to characterize how manifolds can constrain and explain behavioral phenomena in a variety of species and tasks. For example, neural manifolds help explain notable features of grid cells in the mouse entorhinal cortex, as well as head orientation cells in flies, zebrafish and mice.
As the term “neural manifold” has taken root in the field of neuroscience, its meaning has evolved beyond its mathematical origins. In my definition of neural manifolds, there are three key assumptions we should make. First, neural population activity is confined to a surface: There is a geometry to the states that activity can take. Second, this surface is smooth, or differentiable, and continuous, meaning that activity will follow this surface over time. And last, the surface’s intrinsic dimensionality (more on this later) is smaller than the full population dimensionality, meaning that activity states cannot explore the full space. Although this last claim draws a strong link to notions of dimensionality reduction, which find projections or embeddings to simplify neural population data, we should not fully equate that idea with neural manifolds.
Others may disagree with my definition. Indeed, these varying interpretations and definitions lead to several controversies and misconceptions, which I would like to address.
Misconception 1: Neural manifolds are just dimensionality reduction (and vice versa).
Currently, neural manifolds are estimated from neural population recordings post hoc, and the technique used to estimate the neural manifold must be carefully considered. Merely performing dimensionality reduction is not necessarily an appropriate analysis of the neural manifold. One must also consider the properties of the surface, including its topology and geometry (intuitively, its structure and shape). Although it’s easy to conceptualize and visualize these manifolds as flat planes, cartoons such as the one in this article are likely simplified abstractions or approximations of the real manifold geometry in the brain.
Also, given the high degree of recurrence and nonlinearities in neural circuits, it is reasonable to assume that neural manifolds should in fact be nonlinear, as demonstrated by recent work. As such, linear dimensionality reduction—such as principal component analysis (PCA), which identifies hyperplanes in neural space and can, technically, constitute a manifold—may distort the true structure of the data and give an inaccurate or incomplete description. The apparent disconnect between complex nonlinear manifolds and the predominantly linear methods that dominate the literature leads to our next misconception.
Misconception 2: Manifolds are “low dimensional.”
The notion of dimensionality—and the varying definitions scientists use—is ripe for misunderstanding. Phrases such as “low dimensional” and “high dimensional” can be confusing without adequately defining the type of dimensionality. To illustrate, imagine the manifold is a bowl embedded in 3D neural space. Indeed, you cannot fully characterize this bowl without using all three dimensions; by most metrics used in neuroscience, this state space would seem to be three dimensional. Yet, if we imagine flattening the bowl, we can fully describe the neural activity with only two dimensions. I will borrow terminology from Mehrdad Jazayeri and Srdjan Ostojic, who refer to these as the “embedding dimensionality” and “intrinsic dimensionality,” respectively.
In my definition above, the neural manifold framework posits only that the intrinsic dimensionality is smaller than the number of neurons in the population of interest. The full population may be necessary to characterize the neural manifold, meaning that the embedding space may be full dimensional, and recent studies have shown this may be the case. But I predict that the intrinsic dimensionality is significantly lower than the number of neurons because of network recurrence and redundancy—not to mention the inherently low dimensionality of behavior evoked in typical laboratory tasks—and preliminary evidence supports this assumption. A corollary of this prediction is that the functional processes that guide behavior are likely distributed across large populations of neurons in the brain. Yet a common misconception overextends this prediction to mean that individual neurons have nothing to tell us about the brain.
Misconception 3: The manifold dismisses single neurons.
The manifold view and the single neuron view are often presented as a false dichotomy. The importance of individual neurons is well demonstrated, and small numbers of specific neurons can be essential for brain functions such as memory recall. But the brain is a highly recurrent circuit, and the inputs and outputs of neurons are in nearly all cases other neurons, even in peripheral sensory regions that can, in theory, be dominated by the external world. The neural manifold framework need not dismiss an individual neuron’s computations, but it does natively incorporate that the activity of any given neuron is best understood in relation to the activity of other neurons that may provide its inputs—either directly, such as via synapses, or indirectly, such as via neuromodulators. As such, the analysis of neural manifolds and the trajectories along them are an ideal level to help bridge the highly granular activity of individual neurons to the macroscopic processes underlying animal behavior.
Ultimately, the neuron is a fundamental computational unit of the brain. The neural processes that guide animal behavior are, however, in my view, emergent from the biophysical processes that govern each individual neuron. That is, because the processes themselves are specified at the population level, we must observe the population as a whole to describe those processes.
To continue the analogy above (and paraphrase an argument from a conversation with my colleagues Juan Gallego and John Krakauer), you would not summarize the arguments of this article to your colleague by listing the definition of each component word. You would distill higher-level ideas formed by the grammar and organization of those words. I posit that we must consider a similar grammar and high-level structure in neural activity to describe how brains generate the intricacies of animal behavior, and neural manifolds provide a conceptually appealing framework to begin this quest.