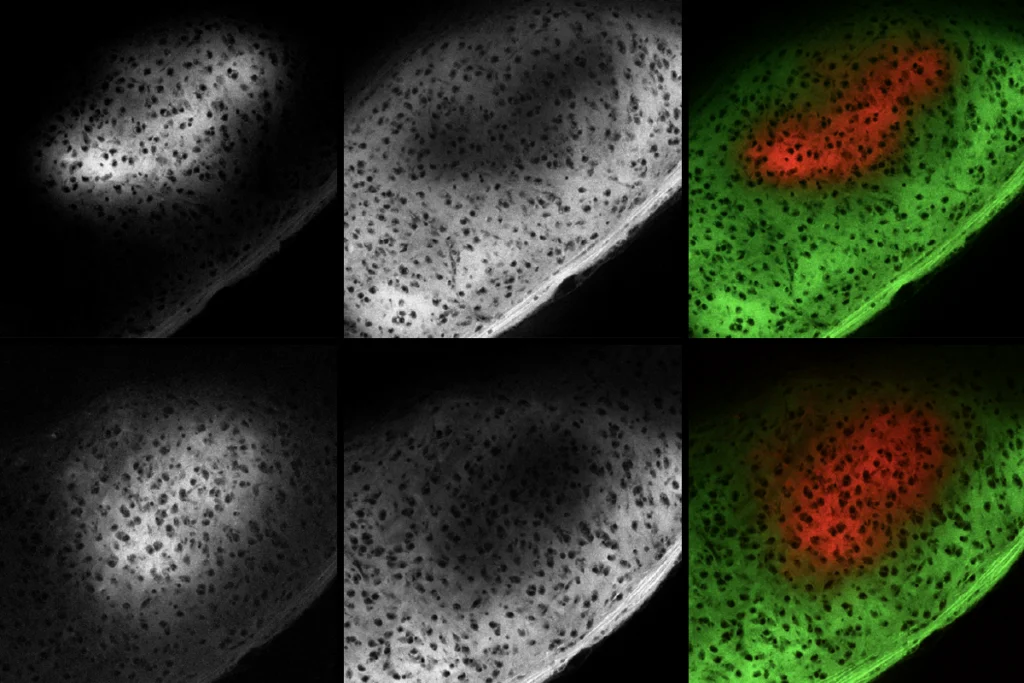
Twenty years ago, a remarkable discovery upended our understanding of the range of elements that can shape neuronal function: A team in Europe demonstrated that enzymatic digestion of the extracellular matrix (ECM)—a latticework of proteins that surrounds all brain cells—could restore plasticity to the visual cortex even after the region’s “critical period” had ended. Other studies followed, showing that ECM digestion could also alter learning in the hippocampus and other brain circuits.
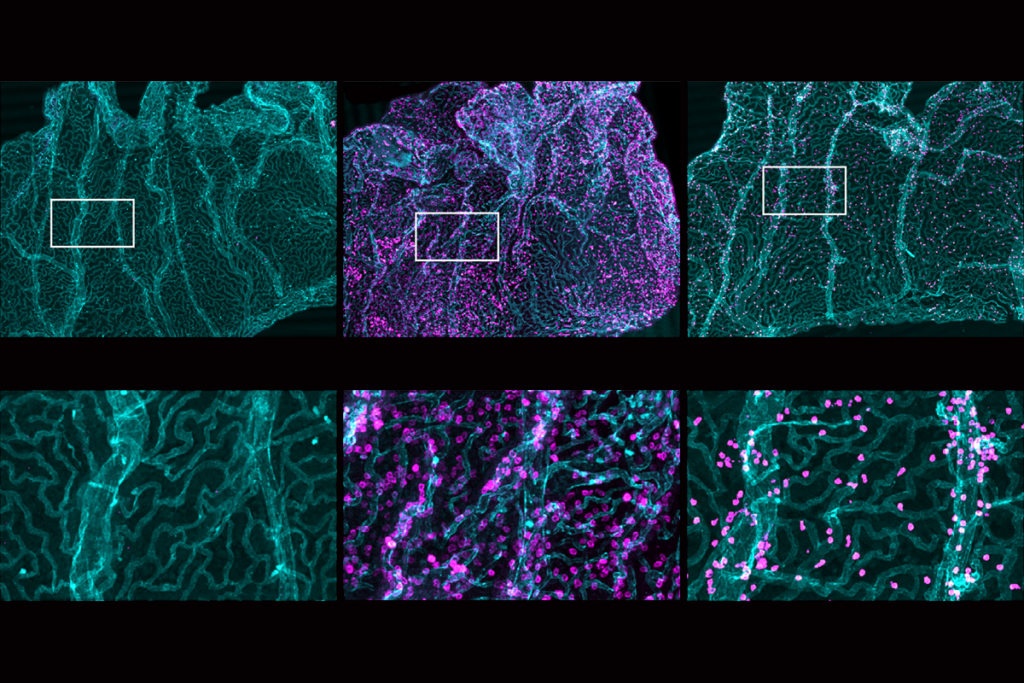
These observations established that proteins outside neurons can control synaptic plasticity. We now know that up to 20 percent of the brain is extracellular space, filled with hundreds of ECM proteins—a “matrisome” that plays multiple roles, including modulating synaptic function and myelin formation.
ECM genes in the human brain are different than those in other species, suggesting that the proteins they encode could be part of what makes our brains unique and keeps them healthy. In a large population study, posted as a preprint on bioRxiv last year, that examined blood protein biomarkers of organ aging, for example, the presence of ECM proteins was most highly correlated with a youthful brain. Matrisome proteins are also dysregulated in astrocytes from people at high risk for Alzheimer’s disease, another study showed.
Despite the influence of these proteins and the ongoing work of a few dedicated researchers, however, the ECM field has not caught on. I would challenge a room full of neuroscientists to name one protein in the extracellular matrix. To this day, the only ECM components most neuroscientists have heard of are “perineuronal nets”—structures that play an important role in stabilizing synapses but make up just a tiny fraction of the matrisome. A respectable scientific journal, covering its own paper that identified a critical impact of ECM, called it “brain goop.”
Why has this area of research languished? I would argue that the field has been hampered by blind spots in our scientific thinking. One of these blind spots is the increasingly challenged assumption that neurons do everything. Aside from oligodendrocytes—which are generally agreed to be important for myelin formation—other non-neuronal cells are still a bit like the unwelcome guests at the holiday table. They complicate things. But these cells are essential to understanding brain function and the ECM.
Research spearheaded by the late Ben Barres established that proteins made by astrocytes can promote synapse formation. A closer look reveals that all of these astrocyte proteins are ECM proteins—they are secreted into the extracellular space and help to build the ECM all over the body. Work from our group shows that microglia promote synapse formation by clearing space in the ECM. I would argue that this activity may be the defining function of microglia in regulating synapse numbers.
These results suggest that a large part of glial biology is the ECM and its effect on neurons. Indeed, many glial biologists have been studying the ECM all along without focusing on it as a field of study. Shifting their perspective could help to advance the field.
P
utting scientific blind spots aside, the ECM field has been similarly held back by technical limitations. Single-cell transcriptomics tools are essential for understanding neuronal and non-neuronal cell function. Yet anyone hoping that transcriptomic approaches are the key to characterizing the ECM is likely to be disappointed. A tremendous amount of regulation happens during protein translation and beyond. Many of the proteins in the ECM are modified by the addition of sugar groups and other changes, which are difficult to predict from the genetic makeup of a cell. We do not yet know if it matters which cell type makes these proteins, or whether the addition of sugar groups alters their functions.To solve the ECM puzzle, we need to focus on proteins, not genes. This will require a systematic mapping of the proteins outside of cells across brain regions, over developmental time, in humans and in model organisms. These efforts should be structured similarly to those applied to gene-expression atlases and brain-connectivity maps, as supported by endeavors such as the BRAIN Initiative.
Fortunately, there have been major advances in proteomics in recent years. This progress will help reveal the importance of the ECM in human disease and enable the mapping of ECM proteins with unprecedented resolution, using strategies such as proximity labeling. But we also must continue to develop technologies to characterize post-translation modifications, such as methods to define how sugar chains are added to proteins and how they modify their function.
But perhaps the most essential step to crack the ECM code is not new technologies but rather a change in thinking. Glial biologists, electrophysiologists and systems neuroscientists need to join forces and consider ways in which we have been studying different facets of the same problem. We need precision, both in how we define the ECM and in the experimental manipulations we use to study it.
It took years of painstaking efforts to define the molecular motors and recognition molecules that govern synaptic vesicle formation, docking and fusion—processes that are now fundamental tenets of neuroscience. Imagine the possibilities for discovery if we understood the connectivity of the matrix outside of synapses at the same level of detail. If the matrix involves the same degree of complexity and precision, the potential payoff for our understanding of brain plasticity and brain diseases from development to degeneration could be immense.